Atlas de Histologia. Cap. 1: Tecido Epitelial de Revestimento

O tecido epitelial está ligado a ações de revestimento em nosso corpo, tanto externo quanto interno. Também está ligado a absorção de nutrientes, isolamento, trocas iônicas, secreção exócrina e endócrina, condução de massas fluidas, etc. Abaixo dos epitélios, existe uma trama feita por proteínas da matriz, a lâmina basal. Ela é composta por um tipo de colágeno (colágeno IV), por um proteoglicano (perlecana), e por duas glicoproteínas, uma delas a laminina. Laminina e fibronectina, outra proteína de ligação de matriz extra-celular com as células epiteliais, formam uma sub-região da lâmina basal denominada "lâmina rara". A malha molecular de colágeno IV (que forma a "lâmina densa") aparenta o formato de uma tela de galinheiro; sua eficiência na filtragem, um dos papéis dos epitélios, é proporcional ao número de "meshs" apresentado, conferindo espessura variável, relacionada às necessidades do epitélio. A laminina forma redes, que garante força tênsil para a lâmina basal. Também a entactina e a proteoglicana perlecana estão presentes na lâmina basal. Entactina é um nidogênio que, como o nome indica, age como um ligante, unindo as malhas de colágeno e laminina. Pode também desempenhar um papel em interações de células com a matriz extracelular. Perlecana é um componente essencial da matriz extracelular dos epitélios vasculares, interagindo com os componentes da matriz e ajudando a manter a função da barreira endotelial.
 |
| Visão esquemática da interação epitélio-lâmina basal-tecido conjuntivo. Disponível em: http://www.ufrgs.br/biologiacelularatlas/fotos/foto_matriz_1a.jpg |
 |
| Aspecto molecular da lâmina basal, em visão esquemática. Disponível em http://www.proteoderm.com/wp1-proteoderm-intro.html |
A
união das células com os componentes da matriz extracelular é
regulada por moléculas de adesão celular, as CAM. Uma das
principais classes de CAM são as integrinas, que ligam a
célula com a fibronectina e laminina, bem como a outras células.
Fibronectinas ligam com a matriz e facilitam sua ligação a
integrinas. A ligação da fibronectina ao domínio extracelular
inicia uma rota de sinalização intracelular, permitindo que o
citoesqueleto auxilie a célula a se adaptar às alterações do
meio.
 |
| visão esquemática dos tipos de junção presentes nos epitélios. Note a interação da malha de microfilamentos de actina com os microvilos e as zônulas de adesão. |
Células
de epitélios são mantidas unidadas por junções. Junções são
estruturas que permitem a integralidade tecidual. Elas são
responsáveis pela manutenção da forma tissular, pela formação de
uma rede funcional entre o citoesqueleto e a matriz extracelular.
Junções são formadas por proteínas integrais da membrana,
unidadas a elementos de citoesqueleto e da matriz extracelular. Entre
os principais tipos de junções encontradas nas células humanas,
destacamos:
Zônula
ocludente ou tight
junctions, são junções nas
quais ocorre íntima associação entre as membranas adjacentes,
formando uma barreira virtualmente impenetrável. Presente apenas em
vertebrados, elas constituem, com os desmossomos e as zônulas
aderentes, os chamados complexos
juncionais. Nos invertebrados, essas
junções correspondem às junções
septadas. Tight
junctions são compostas por uma
rede ramificada, de pontos selados, extremamente eficientes para
bloquear a passagem de íons. Cada ponto selado é formado por um
agregado de proteínas transmembrana, embebidas em ambas membranas
plasmáticas, como se fosse um ponto de bordado. Embora várias
outras proteínas tenham sido descritas para as zônulas ocludentes,
os principais tipos de proteínas seladoras dessas junções são as
claudinas
e ocludinas.
Essas proteínas associam-se com filamentos de actina, unindo, desta
forma, o citoesqueleto de células adjacentes.
As
junções ocludentes apresentam várias funções:
- Manter as células unidas;
- Bloquear o movimento de proteínas integrais transmembrana entre a superfície apical e basolateral, garantindo a manutenção de domínios de membrana especializados.
- Previne a passagem de moléculas e íons pelo espaço intercelular. Na barreira hemato-encefálica, esse papel é crítico para o isolamento do tecido nervoso.
Entre
as principais proteínas da ZO, estão:
- Zonulina: é o alvo principal da toxina do vibrião da cólera, Vibrio cholerae. Também está implicada na patogênese da doença celiaca e do diabetes mélito tipo 1.
- Claudinas: são proteínas importantíssimas para a constituição das zônulas ocludentes. Elas estabilizam a barreira paracelular que controla o fluxo de moléculas no espaço intercelular
 | |
| visão esquemática de desmossomos, zônulas de oclusão e junções Gap. |
Os
desmossomos, ou macula adherens,
são estruturas especializadas na adesão celular, cuja morfologia
permite que ajude a célula a resistir a forças de tensão e outras,
exercida sobre as mesmas. Nessas junções, o espaço intercelular é
bastante amplo (30 nm). Desmossomos são encontrados unindo células
epiteliais e musculares. Nos desmossomos, existe a ligação entre
proteínas de adesão da membrana plasmática com elementos do
citoesqueleto, notadamente a citoqueratina. Hemidesmossomos são junções que unem a membrana plasmática da célula com a lâmina basal.

Proteínas
de adesão celular do desmossomo, a desmogleína
e a desmocolina,
pertencem à famíla da caderina de moléculas de adesão celular.
Elas são proteínas transmembrana, que ligam à célula epitelial
adjacente por meio de ligação homofílica de seus domínios
extracelulares com outras caderinas de outros desmossomos de células
adjacentes. Ambos possuem cinco domínios extracelulares, e
apresentam seqüências peptídicas com alta afinidade ao cálcio.
O
domínio extracelular do desmossomo é denominado de desmogléia,
e é cortada por uma linha média eletrondensa, onde a desmogleína e
desmocolina ligam-se uma à outra. Essas proteínas podem ligar em W,
S ou λ.
 |
| visão esquemática de um desmossomo |
No
lado citoplasmático da membrana plasmática,existem duas estruturas
densas chamadas de placa densa
externa, e uma placa
densa interna. A placa densa externa
é onde os domínios citoplasmáticos das caderinas ligam-se na
desmoplaquina, via placoglobina e
placofilina. A placa densa interna é
onde os filamentos intermediários ligam no desmossomo. Devido à presença de desmossomos em abundância, um dos estratos da pele é denominado estrato espinhoso.
 |
| estrato espinhoso, pele fina. Notar, na periferia das células, os pontos onde ocorrem os desmossomos. |
Em
algumas patologias autoimunes, como o pênfigo bolhoso, ocorre
defeitos nos desmossomos, que dão aspecto anômalo à pele.
Pacientes com pênfigo desenvolvem anticorpos contra as caderinas.
Zônula
aderente
São
complexos protéicos que ocorrem em junções mais basais que as
junções íntimas. Tais junções têm aspecto de uma faixa
circulando internamente a célula. Uma junção similar, que ocorre
em células não-epiteliais, é a fáscia
aderente. Sua estrutura é a mesma,
mas o padrão de faixa ao redor da célula não fecha completamente o
perímetro celular. Um exemplo é o cardiomiócito. Essa junção é
composta de três proteínas:
- Caderinas: proteínas transmembrana que formam um homodímero cálcio-dependente.
- Beta e alfa cateninas: formam um complexlo com a porção intracelular da caderina.
As
junções aderentes foram, por muitos anos, ligadas à função de
ancorar células, empregando os microfilamentos de actina. O modelo
aceito hoje é de que esta junção atua como ponte, conectando o
citoesqueleto de actina de células vizinhas, através de interação
direta. A zônula aderente pode servir como módulo regulatório para
manter o anel contrátil de actina, o que lhe é imputado em estudos
ao microscópio.

- Permitir a comunicação elétrica direta entre as células, a partir dos conéxons;
- Permitir comunicação química entre as células, viabilizando a passagem de vários segundos mensageiros, com inositol fosfato, cálcio e AMPc.
- Permitir a passagem de moléculas menores que 1000 Da.
Novas
funções têm sido descritas para as junções gap: a) acoplamento
elétrico e metabólico entre as células; b) Trocas elétricas e
metabólicas através dos hemicanais; c) Permitir a ação de Genes
Supressores de Tumor (Cx43, Cx32 e Cx36) d) Função adesiva
independente da junção condutiva.
No
coração, as junções gap auxiliam na conexão dos cardiomiócitos,
dando o aspecto de “sincício funcional” ao miocárdio. Em
neurônios do núcleo vestibular, no núcleo do nervo trigêmeo, no
núcleo olivar inferior e na área tegmental ventral, ocorre o
acoplamento elétrico através de junções gap. No locus
ceruleus, foi observado um fraco
acoplamento de células da glia a neurônios, e no cerebelo,
entre as células de Purkinje
e a glia de Bergmann.
Alguns autores afirmam que astrócitos unem-se fortemente por junções
gap. Dados experimentais dão conta da forte expressão das gaps
nessa classe de células gliais.
Para
que ocorram as junções, uma classe especial de proteínas, as
moléculas de adesão celular,
ou CAMs,
promovem a ligação das células umas com as outras, ou destas com
os elementos da matriz extracelular. Essas proteínas atuam como
receptores transmembrana, sendo compostos por três domínios: um
domínio intracelular, que interage com o citoesqueleto; um domínio
transmembrana; e um domínio extracelular, que interage com as CAMs
da mesma espécie (ligação
homofílica) ou com a matriz
extracelular (ligação
heterofílica). Destacamos as
caderinas, cateninas, selectinas, integrinas e desintegrinas, como
principais grupos de proteínas de adesão celular.
Sobre a forma e função de epitélios, em geral, podemos classificá-los em glandulares (predominantemente secreção endócrina, exócrina ou ambas) e de revestimento (função primordial de revestimento e ações afins). Este último grupo pode ainda ser classificado de acordo com o número de camadas celulares: simples (camada única de células), estratificado (duas ou mais camadas de células), pseudo-estratificado (falso aspecto de estratificação, na realidade é monocamada) e de transição (hoje em dia é considerada uma variante do pseudo-estratificado, com células que modificam sua morfologia conforme o estado funcional do órgão). Ainda conforme a morfologia das células que compõe a camada mais externa (ou a única, no caso dos epitélios simples), podemos classificar os tecidos epiteliais de revestimento em pavimentosos/escamosos (células achatadas, núcleo acompanhando a morfologia, similares a escamas de peixe ou azulejos), cúbicos (formato cuboidal) ou cilíndrico/prismático/colunar (formato cilíndrico, com núcleo orientado no eixo maior de célula).
Entre as mais notáveis
estruturas celulares ligadas aos epitélios, especificamente nas vias aéreas e tuba uterina, bem como aos espermatozóides, estão os cílios
e flagelos.
Também há cílios associados às células ependimárias, que, no entanto, não são consideradas um verdadeiro epitélio, devido à falta de uma lâmina basal associada. Cílios e flagelos são estruturas ligadas a movimento, tanto da
própria célula quanto de massas fluídas. São os responsáveis
pela propulsão do espermatozóide, de vários protozoários
flagelados e ciliados, pela funcionalidade do elevador mucociliar do
trato respiratório, pela movimentação do líquido
céfalo-raquidiano e do óvulo fecundado ao longo da tuba uterina.
Um cílio se compõe de
um grupo de microtúbulos recoberto por membrana plasmática,
recebendo a denominação de axonema.
Cada axonema apresenta um anel de nove microtúbulos duplos, que
rodeiam o par central de microtúbulos simples. Cada dupla exterior é
composta de um anel de 13 filamentos, ou subfibra A, fundidos a outro
conjunto de 10 filamentos, a subfibra B. Os filamentos dos
microtúbulos são compostos de heterodímeros
de alfa e beta tubulina.
Os 11 microtúbulos que compõem o axonema se mantém unidos por três
tipos de conectores: as díades exteriores adjascentes estão unidas
por uma proteína elástica denominada nexina.
Nas subfibras A estão os braços
de dineína,
externos e internos. A dineína, por ação ATPásica, é a
responsável pelo movimento dos flagelos e cílios. As díades de
microtúbulos estão unidas ao par central de microtúbulos por
pontes
radiais.
Por sua vez, o par central está circundado por uma bainha
central.
Os cílios podem ser classificados de duas maneiras. Acima, nos referíamos aos chamados cílios motores, típicos dos epitélios que requerem movimento de massas fluídas, e que são essenciais para o funcionamento do trato respiratório, do sistema ventricular do encéfalo e na condução do zigoto ao útero. Contudo, ocorrem cílios associados à captação de estímulos, tendo, portanto, função sensorial. São os chamados cílios primários. Sua função, portanto, é a de ser uma espécie de órgão sensorial estabilizado dinamicamente por microtúbulos. Eles representam papéis importantes na mecano e químio-percepção, e suas disfunções estão ligadas a desordens no desenvolvimento e muitas doenças.
Os cílios podem ser classificados de duas maneiras. Acima, nos referíamos aos chamados cílios motores, típicos dos epitélios que requerem movimento de massas fluídas, e que são essenciais para o funcionamento do trato respiratório, do sistema ventricular do encéfalo e na condução do zigoto ao útero. Contudo, ocorrem cílios associados à captação de estímulos, tendo, portanto, função sensorial. São os chamados cílios primários. Sua função, portanto, é a de ser uma espécie de órgão sensorial estabilizado dinamicamente por microtúbulos. Eles representam papéis importantes na mecano e químio-percepção, e suas disfunções estão ligadas a desordens no desenvolvimento e muitas doenças.
O corpúsculo basal,
que está ligado à montagem dos cílios primários, deriva dos
centríolos maduros, que por sua vez originam-se dos centrossomos,
como vimos. Muias proteínas centrossomais, portanto, estão
implicadas na organização ciliar. Uma proteína identificada há
pouco tempo, a Cep164, é indispensável à ciliogênese, epode ser
localizada nas porções distais dos centríolos maduros. Em
contraste com outras proteínas do centrossomo, como a nineína e a
Cep170, que são componentes dos apêndices subdistais, Cep164
persiste em centríolos durante a mitose. A localização de Cep164 e
nineína/Cep170 demonstra-se mutualmente independente durante a
interfase. Portanto, a Cep164 é um excelente marcador das porções
distais de cílios maduros.
Outra projeção celular importante nos epitélios são as microvilosidades ou microvilos. As
microvilosidades ou microvilos são envoltas por membrana cujo interior é formado de
25 a 30 filamentos de actina, ou microfilamentos. Apresentam variações em sua
aparência, de modo que, em algumas células, as microvilosidades são
projeções curtas, irregulares e bolhosas, em outros tipos
celulares, são projeções altas, uniformes e muito próximas, por
vezes de formato mais alongado, lembrando cílios imóveis (daí o
nome de estereocílios para as projeções do epidídimo),
as quais aumentam muito a área da superfície livre celular. O
número e o formato de
microvilos de um determinado tipo celular apresentam relação
estreita com sua função.
Estas projeções são sustentadas por microfilamentos. Sua ocorrência é predominantemente apical nas células epiteliais, mas podem, eventualmente, ocorrer nas regiões laterais de células polarizadas. Microvilos ampliam a superfície da membrana plasmática aumentando sua eficiência para as trocas com a cavidade ou o meio extracelular. Os microfilamentos de actina e proteínas associadas preenchem e sustentam tais especializações, penetrando profundamente no citoplasma, na base das projeções, interagindo com os demais elementos do citoesqueleto na região apical da célula. Essa concentração de citoesqueleto na base das projeções, denominada trama terminal, é facilmente observada ao microscópio de luz como uma linha fortemente corada.
Estas projeções são sustentadas por microfilamentos. Sua ocorrência é predominantemente apical nas células epiteliais, mas podem, eventualmente, ocorrer nas regiões laterais de células polarizadas. Microvilos ampliam a superfície da membrana plasmática aumentando sua eficiência para as trocas com a cavidade ou o meio extracelular. Os microfilamentos de actina e proteínas associadas preenchem e sustentam tais especializações, penetrando profundamente no citoplasma, na base das projeções, interagindo com os demais elementos do citoesqueleto na região apical da célula. Essa concentração de citoesqueleto na base das projeções, denominada trama terminal, é facilmente observada ao microscópio de luz como uma linha fortemente corada.
No
tocante à organização morfológica, as microvilosidades do
epitélio intestinal são mais ordenadas e uniformes na aparência
que aquelas que constituem a borda em escova das células renais. O
nome de “borda
estriada”
é
constantemente atribuído à margem com microvilos dos enterócitos;
na superfície
das células que revestem os túbulos contorcidos proximais do
néfron, na
córtex do
rim, utiliza-se
o termo “borda
em escova”.
Nos demais tecidos sua ocorrência pode ser em menor número e por
vezes com comprimento variável. Apresentam um núcleo de filamentos
de actina ancorados na vilina, localizada no ápice da
microvilosidade, e que se estendem para baixo e para dentro do
citoplasma apical. Há uma interação com uma rede horizontal de
filamentos de actina, a trama terminal, que se localiza abaixo da
base das microvilosidades, estabilizada através da espectrina, que
também fixa a trama terminal à membrana celular apical. A trama
terminal apresenta capacidade contrátil e esta é devida à presença
da miosina II e da tropomiosina.
 |
| epitélio intestinal: observar o bordo em escova (microvilosidades) |
Sobre a forma e função de epitélios, em geral, podemos classificá-los em glandulares (predominantemente secreção endócrina, exócrina ou ambas) e de revestimento (função primordial de revestimento e ações afins). Este último grupo pode ainda ser classificado de acordo com o número de camadas celulares: simples (camada única de células), estratificado (duas ou mais camadas de células), pseudo-estratificado (falso aspecto de estratificação, na realidade é monocamada) e de transição (hoje em dia é considerada uma variante do pseudo-estratificado, com células que modificam sua morfologia conforme o estado funcional do órgão). Ainda conforme a morfologia das células que compõe a camada mais externa (ou a única, no caso dos epitélios simples), podemos classificar os tecidos epiteliais de revestimento em pavimentosos/escamosos (células achatadas, núcleo acompanhando a morfologia, similares a escamas de peixe ou azulejos), cúbicos (formato cuboidal) ou cilíndrico/prismático/colunar (formato cilíndrico, com núcleo orientado no eixo maior de célula).
Exemplificando:

Rim, córtex, coloração: PAS/Hematoxilina. Observe a cápsula de Bowman, delineada em rosa-choque. É um típico caso de epitélio pavimentoso simples. Os túbulos renais, amplamente vistos na imagem, são exemplos de tecido epitelial cúbico simples.

Detalhe em grande aumento dos túbulos contorcidos distais (seta) e proximal, na córtex renal, corados com azul de toluidina. Ambos são epitélios cúbicos simples, em que pese a diferença na altura e morfologia das células dos dois tipos de túbulos nefrônicos. A presença de microvilos nos túbulos proximais revela sua função na reabsorção de materiais presentes no ultra-filtrado glomerular. Os microvilos de túbulos contorcidos proximais renais formam a chamada "borda em escova". Junto a eles, pode ser vista a trama terminal, em sua base, fortemente corada. Note ainda o labirinto basal, intensamente corado de azul, na base das células cúbicas do TCP. O labirinto basal é um tipo de invaginação da membrana, que permite o alojamento das mitocôndrias em paliçada típicas dessas células.

Duodeno, HE. Note o tecido epitelial simples cilíndrico, com suas células altas, dotadas de microvilosidades absortivas (bordo estriado). Também percebemos as células caliciformes, aqui vistas com aspecto globoso e branco. Essas células secretam muco, essencial para o trânsito intestinal. Associado ao epitélio está o tecido conjuntivo frouxo, que forma a lâmina própria da mucosa intestinal.

Detalhe do epitélio acima, mostrando várias células caliciformes.

Epitélio pseuto-estratificado ciliado cilíndrico da traquéia. Note os cílios, essenciais para movimentação do muco produzido nas vias aéreas, entre outras áreas do nosso corpo.

Esôfago: epitélio pavimentoso estratificado. Note que este epitélio apresenta várias camadas de células. Este tipo de epitélio pode ser visto ainda na boca, ânus e vagina, ou seja, superfícies úmidas do corpo.

Pele espessa: epitélio pavimentoso estratificado queratinizado. Diferente do epitélio acima, este tipo de tecido epitelial está presente na pele, ou seja, em superfície seca externa. A camada mais externa, ou estrato córneo, é formada essencialmente por células anucleadas, mortas, o que protege mecanicamente as camadas inferiores.

Epitélio cilíndrico estratificado. Presente na uretra peniana.

Epitélio cúbico estratificado. Presente em ductos de glândulas salivares e sudoríparas.
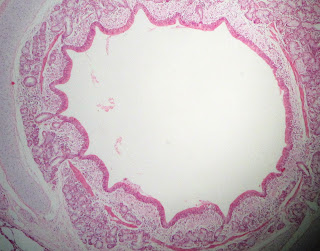
Epitélio de transição ou urotélio. Como o nome sugere, está presente em vias urinárias, como na pelve renal, ureteres e bexiga.

Comentários
Postar um comentário